
A comprehensive review of loading controls for Western blots, and the survey results from formal publications.
蛋白质印迹用于测量不同条件下特定蛋白的变化。在蛋白质印迹过程中涉及许多步骤,包括样品制备、电泳、蛋白转移、抗体孵育和信号检测。为了解释蛋白质印迹实验的结果,一个贯穿全过程的内参是必须的。
内参保证了
- 每个泳道中加入了相同量的蛋白样品,
- 不同泳道上的蛋白以相同的效率从凝胶转到膜上,
- 不同泳道上抗体孵育(对于一抗而言,必要的话也包括二抗)、信号检测都是一致的。

内参的信号通常用来对目标蛋白的信号进行标准化。需要强调的是,对照蛋白抗体应该与实验抗体在相同的印迹上进行。
来邦网调研了关于蛋白质印迹中内参的文章。结果表明肌动蛋白(特别是β肌动蛋白)是最常用的参照。图一展示了从部分调研文献中整理出来的蛋白质印迹图片 [1, 2, 2-5] 。此次调研的其它有趣结果会在本文的稍后部分进行讨论。
| 内参 | 供应商 | 引用次数 | 代表性文献 |
|---|---|---|---|
| 肌动蛋白 | 56 | ||
| Sigma-Aldrich | 30 | ||
| Santa Cruz Biotechnology | 12 | ||
| EMD Millipore/Chemicon | 3 | ||
| Abcam | 2 | ||
| Roche | 2 | ||
| MP Biomedicals | 1 | 691001 [6] | |
| 微管蛋白 | 21 | ||
| Sigma-Aldrich | 10 | ||
| Santa Cruz Biotechnology | 5 | ||
| MilliporeSigma | 1 | CP06 [6] | |
为内参的蛋白必须要符合一定的标准。这些标准是:
- 在测试条件下内参的蛋白水平要保持恒定(与总蛋白量相比),例如,用特定药物处理或未经特定药物处理,或在发育阶段期间,
- 内参的检测带与那些目标蛋白的检测带在分子量上有所不同,
- 内参和目标蛋白的检测限都在动态范围内。也就是说,这种检测方法和操作流程可以在信号不饱和的情况下揭示内参和目标蛋白量的变化。
对于特定的蛋白质印迹操作流程而言,一个参照的动态检测范围可以通过针对性地对一系列稀释度的该蛋白对照进行蛋白质免疫印迹并检测信号来确定。信号强度必须和稀释度相关联。
不同的样品制备需要不同的内参。表二总结了针对不同样品制备常用的内参,包括全细胞/胞浆蛋白、线粒体、核蛋白、植物组织和血清样本。下面将一一详细讨论。
| 样品 | 内参 | 分子量 |
|---|---|---|
| 全细胞/胞浆蛋白 | ||
| β-肌动蛋白 | 43 | |
| α-肌动蛋白 | 43 | |
| GAPDH | 30-40 | |
| β-微管蛋白 | 55 | |
| α-微管蛋白 | 55 | |
| (高分子量) | 黏着斑蛋白 | 116 |
| 线粒体 | ||
| VDCA1/孔蛋白(porin) | 31 | |
| 细胞色素C氧化酶(cytochrome C oxidase) | 16 | |
| 核蛋白 | ||
| 核纤层蛋白B1(lamin B1) | 66 | |
| TATA结合蛋白(TATA binding protein TBP) | 38 | |
| PCNA | 29 | |
| 组蛋白 H1 | 30 | |
| 组蛋白 H3 | 18 | |
| 植物组织 | ||
| LHCP | 25 | |
| APX3 | 32 | |
| 血清 | ||
| 转铁蛋白 | 77 | |
| 肌肉 | ||
| SDHA [7] | 73 | |
| 酵母 | ||
| 磷酸甘油酯激酶 | 45 | |
在人和其它脊椎动物中,肌动蛋白是由六种蛋白组成的蛋白家族,共分为3组: alpha ( ACTC1, cardiac muscle 1), alpha 1 ( ACTA1, 骨骼肌) and 2 ( ACTA2, 主动脉平滑肌), beta ( ACTB), gamma 1 ( ACTG1) and 2 ( ACTG2, 肠道平滑肌)。beta和gamma 1是两种非肌型肌动蛋白。它们充当微丝的主要组成部分。肌动蛋白是高度保守的。来源于各种如人、小鼠和鸡的beta肌动蛋白都是相同的。人源beta肌动蛋白与真菌的同源蛋白有近90%的相同性。人源beta肌动蛋白与人源肌动蛋白家族中的其它成员至少有93%是相同的。这些肌动蛋白家族成员间以及种族间的同源性信息对于抗体选择和解释蛋白质印迹条带是很重要的。
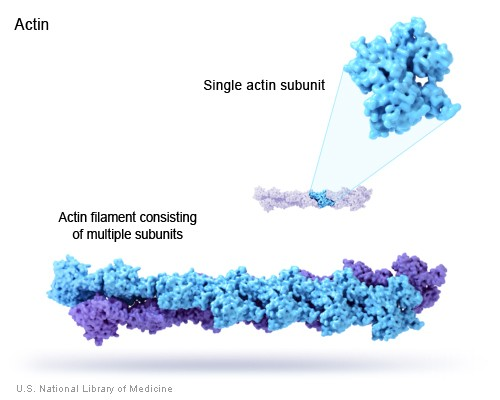
beta和alpha肌动蛋白在蛋白质印迹实验中一直被用作内参。 请参阅有关beta肌动蛋白内参、常用克隆和beta肌动蛋白对照常见问题的详细讨论。肌动蛋白在细胞中浓度往往是很高的,因此为了检测到肌动蛋白量的任何变化,调整上样体积和蛋白质印迹操作流程是很重要的。 细胞生长条件的改变会破坏肌动蛋白的合成。有报道表明beta肌动蛋白在蛋白质印迹分析中可能不是一个可靠的内参 [8] 。beta肌动蛋白也存在于核内,作为染色质重塑复合物的一个组成部分 [9] ,但不能用于核蛋白样品的对照。
糖酵解中 甘油醛-3-磷酸脱氢酶 (GAPDH, G3PD, GAPD)在有无机磷酸盐和烟酰胺腺嘌呤二核苷酸(NAD)存在的情况下催化甘油醛-3-磷酸的可逆氧化磷酸化。它也有亚硝化酶活性并参与基因转录、RNA运输、DNA复制和凋亡。
GAPDH是一个管家基因,在蛋白质印迹和PCR中都被用作参照。GAPDH在物种间是高度保守的。例如,人源由335个氨基酸组成的GAPDH其全长有70%与Arabidopsis thaliana中由422个氨基酸组成的同源蛋白是相同的。 它存在于胞浆、细胞核、核周区和膜上。 一项全面的研究表明GAPDH mRNA水平在不同组织类型中有显著差异,但是相对于年龄和性别而言却是保持恒定的 [10] 。缺氧可上调GAPDH的表达 [11, 12] ,导致GAPDH聚集 [13] 。因此不能在涉及氧气的相关研究中作为内参。有趣的是,它是用于治疗牛皮癣和多发性硬化症的免疫调节药物的靶点,比如富马酸二甲酯,它可以琥珀酸化和灭活GAPDH的催化半胱氨酸 [14] 。
一个侧面说明:GAPDH在特定条件下是与微丝(beta肌动蛋白)相关联的 [15] 。
有五种微管蛋白:alpha、beta、gamma、delta和epsilon。alpha和beta微管蛋白组成的异源二聚体是微管的构成组分。
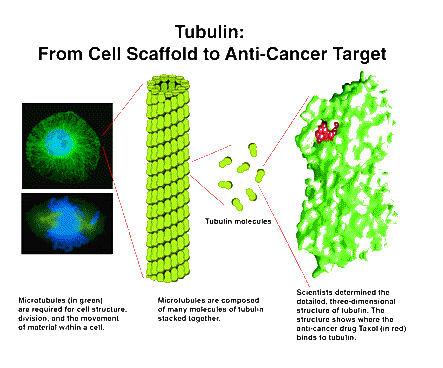
alpha微管蛋白的亚型包括1a (TUBA1A)、1b (TUBA1B)、1c (TUBA1C)、3c (TUBA3C)、3d (TUBA3D)、3e (TUBA3E)、4a (TUBA4A)和8 (TUBA8)。I类(TUBB)、IIa类(TUBB2A)、IIb类(TUBB2B)、III类(TUBB3)、IVa类(TUBB4A)、IVb类(TUBB4B)、V类(TUBB6)、VI类(TUBB1)和VIII类(TUBB8)都是beta微管蛋白的亚型。
gamma微管蛋白,gamma 1(TUBG1)和2(TUBG2),参与微管的成核和极性排列,并存在于中心体和纺锤体极体中。delta(TUBD1)和epsilon(TUBE1)微管蛋白往往位于中心粒处,并且是有丝分裂纺锤体的结构部件。
微管蛋白在物种间是高度保守的。例如,人源有451个氨基酸的微管蛋白alpha 1a与酵母中由445个氨基酸组成的同源蛋白Tub3p有74%是相同的。alpha、beta和gamma的亚型也都是非常类似的。人源alpha微管蛋白相互间有超过90%的相同性。不同型的微管蛋白也有显著的同源性。人源alpha微管蛋白和人源beta微管蛋白有40%相同性。
alpha、beta和gamma微管蛋白都一直被用作内参。
微管蛋白是多组药物的药理靶点,像抗癌药物紫杉醇(Taxol)、Tesetaxel、长春碱(vinblastine)和 长春新碱(vincristine),抗痛风剂秋水仙碱(colchicine)和抗真菌药物灰黄霉素(griseofulvin)。这些药物都会影响体外和体内的微管蛋白表达。
黏着斑蛋白,另一种细胞骨架蛋白,也被用作内参 [16-19] ]。它是细胞与细胞、细胞与基质连接的主要成分之一。人黏着斑蛋白的分子量为117kDa,含有1066个氨基酸。 它可以用作高分子量蛋白质的内参。 在肌肉组织中,还表达了一个含有150KDa分子量的额外外显子的剪接变体。
核纤层蛋白是核纤层的结构部件,用于支撑核被膜并参与细胞周期中核被膜的解体和再形成。在动物细胞中,3种基因编码了至少7种核纤层蛋白。LMNA基因通过可变剪接编码A型和C型核纤层蛋白,而LMNB1和LMNB2分别编码核纤层蛋白B1和B2。B型核纤层蛋白存在于每个细胞。A型核纤层蛋白在原肠胚形成后表达,C型核纤层蛋白的表达是组织特异性的。
核纤层蛋白会进行法尼基化和磷酸化等翻译后修饰,从而调节核纤层的组装和核纤层蛋白的活性。
在核纤层蛋白中,核纤层蛋白B1往往被用作内参。核纤层蛋白B1在物种间是高度保守的。人源核纤层蛋白B1与啮齿目动物中的同源蛋白有95%的相同性,与果蝇的同源蛋白有36%的相同性。人源核纤层蛋白B1与其它人源核纤层蛋白有50%以上的相同性。 核纤层蛋白B1对于没有核被膜的样品是不适合作为内参的。
TATA结合蛋白,TBP,是在由RNA聚合酶II进行基因转录前能特异性结合到TATA框DNA序列的通用转录因子。TATA框存在于10-20%的人类启动子中。TBP的N端有一长串谷氨酰胺(TBP mRNA中的CAG重复),可以调节C端的DNA结合活性。正常人群中TBP N端的谷氨酰胺数量是不尽相同的(25至42个重复),并且在某些病理状态下会延长至47-63个重复。TBP是高度保守的。人源TBP与啮齿目动物中的同源蛋白有90%的相同性,与真菌的同源蛋白有超过60-70%的相同性。
增殖细胞核抗原,PCNA,是DNA聚合酶delta的一个辅助因子,并且在细胞周期的DNA合成阶段(S期)在细胞核内表达。PCNA参与真核DNA复制。PCNA在黑猩猩、狗、牛、大鼠、鸡、斑马鱼、果蝇、蚊子、裂殖酵母、芽殖酵母、克鲁维酵母(K. lactis)、棉阿舒囊霉(E. gossypii)、稻瘟病菌(M. grisea)、粗糙脉孢菌(N. crassa)、拟南芥和水稻中都是保守的。人源由261个氨基酸组成的PCNA与啮齿目动物中的同源蛋白有97%的相同性,与酵母中的同源蛋白有36%的相同性。
PCNA对于非增殖细胞或经过抗增殖处理的细胞并不是一个好的内参。
电压依赖性阴离子通道,作为一类孔蛋白家族,构成了真核细胞中线粒体外膜上的主要蛋白。在脊椎动物中至少有三种成员(VDAC1、VDAC2和VDAC3)。VDAC1在线粒体外膜和质膜上都有,发挥着不同的功能。它表达于心脏、肝和肌肉组织中。由于可变剪接导致了有多种变体。VDAC1基因在黑猩猩、狗、牛、小鼠、大鼠、鸡和斑马鱼中是保守的。人源VDAC1与小鼠的同源蛋白有99%的相同性,与斑马鱼的同源蛋白有85%的相同性。相关例子见 [20] 。
位于线粒体内膜的细胞色素C氧化酶是线粒体电子传递链中的最后一个酶复合体。该复合体由13个线粒体编码和核编码的亚基组成。
细胞色素C氧化酶的亚基IV通常被用作内参。它有两种亚型(亚型1和2)。它们由两个核基因编码(COX4I1和COX4I2)。亚型1是广泛表达的,亚型2是高度表达于肺部组织的。这两种亚型在人当中有60%的相同性。亚型I COX4I1被用作内参。人源COX4I1与小鼠的同源蛋白有80%的相同性,与斑马鱼的同源蛋白有63%是相同的。
转铁蛋白是一种血浆糖蛋白,其主要功能是将铁从肝脏,肠和网状内皮系统转运到全身的增殖细胞。人转铁蛋白具有698个氨基酸,与其小鼠直系同源体具有73%的同一性,与斑马鱼直系同源体具有43%的同一性。 转铁蛋白的表达受一些遗传性疾病和维甲酸治疗的影响。
APX3,抗坏血酸过氧化物酶3,已被用作植物膜蛋白内参。它是一种微粒体抗坏血酸过氧化物酶,可清除植物细胞中的过氧化氢。在拟南芥中:有三种胞质型(APX1,APX2,APX6),两种叶绿体类型(基质sAPX,类囊体tAPX)和三种微粒体型(APX3,APX4,APX5)亚型。
有些作者建议在蛋白质印迹中放弃使用一种蛋白作为内参,并建议要依赖于在转移步骤之前(通过考马斯亮蓝 [21] )或之后(通过丽春红 [22] )对蛋白进行染料染色,或者使用无需染色的技术(在凝胶中加入58-Da 三卤化合物 [23] )。然而这一措施没能够顾及到作为一个理想参照其所有三方面的考量:样品承载、蛋白转移和抗体孵育/信号检测。
因此应该考虑将这三种方式进行组合,即一个蛋白内参、对凝胶的考马斯亮蓝染色和对转移膜的丽春红染色,从构成一个精心设计的蛋白质印迹操作流程。
在来邦网调研的112篇发表文章中,许多供应商的抗体已经被用于检测不同类型的内参。
Santa Cruz的羊源多克隆抗肌动蛋白抗体被用于研究类共济失调毛细血管扩张症和Nijmegen破损综合症中DNA损伤的信号转导 [18] ,以及关节软骨细胞中的研究 [19] 。它的兔源多克隆抗肌动蛋白抗体被用于猪肾小管上皮LLC-PK1细胞的实验 [21] 。Santa Cruz Biotechnology的抗beta肌动蛋白抗体在人脐静脉内皮细胞样品中 [22] ,在小鼠心肌细胞中分离到的核蛋白提取物中 [24] ,在小鼠横纹肌肉瘤细胞和RMS772细胞中 [25] ,在细胞提取物中 [26] ,在突变的和正常的B淋巴细胞中 [27] ,在293T细胞中 [28] 以及视网膜色素上皮细胞中 [29] 都被用作内参。
来自于 EMD Millipore/Chemicon的肌动蛋白抗体被用于从注射过LiCl、载体或alpha MPT的DAT-KO小鼠 [30] 、经NMT1-1 siRNA或NMT2-4 siRNA处理过的HT-29和SK-OV-3细胞制备得到的裂解样品中 [31] 。
Abcam的抗肌动蛋白抗体被用于研究人类巨噬细胞中的ABCA1和ABCG1 [32] ,并被用于786-O、UOK121和UMRC6肾细胞癌细胞中 [33] 。
为了研究Brachyury在肿瘤细胞上皮间质转化和肿瘤恶化中的作用, Thermo Fisher Pierce / Neomarkers的抗beta肌动蛋白被用作内参 [34] 。
为了检测p53、WAF1和MDM2 [35] ,在Hela、野生型H2AX(H2AXflox/flox)和H2AX缺失型胚胎干细胞中 [36] ,在一个B淋巴细胞前体细胞株中 [37] ,为了在猕猴中检测TRIM5蛋白 [38] ,在U937细胞中 [39] ,在成纤维细胞和手术脑样本中 [40] ,在小鼠骨髓细胞中 [41] 以及迁移性T细胞中 [42] ,Sigma的抗肌动蛋白均被用于蛋白质印迹。在转染有PKD RNAi、Hsp27 RNAi和载体对照的HeLa细胞中,Sigma的抗alpha肌动蛋白抗体作为内参用于蛋白质印迹 [43] 。为了确定可能对哺乳动物神经细胞分化极其重要的转录因子 [44] ,在肝细胞中 [45] ,在人类冠状动脉内皮细胞中 [46] ,在转染有Luc siRNA或NudC siRNA的HeLa细胞中 [47] ,在NCI-H460和NCI-H1299细胞中 [48] ,在人星形细胞瘤细胞株1321N1中 [49] ,在TAK1+/+和TAK1-/-小鼠胚胎成纤维细胞和HeLa S3细胞中 [50] ,在HeLa细胞的核及胞浆中 [51] ,在野生型Jurkat细胞和稳定转染有Bcl-xL、Bcl-2的Jurkat细胞或载体对照细胞中 [52] ,在来自于p85缺失小鼠和相应的野生型小鼠的脾B淋巴细胞中 [53] ,在小鼠的胶原蛋白黏附性FN-/-细胞中 [54] 以及在K562-R、LAMA-R和其经1 M甲磺酸伊马替尼处理过的母细胞中 [55] ,都使用了Sigma的抗beta肌动蛋白抗体。
Roche Molecular Biochemicals的抗肌动蛋白(C4)抗体被用于文献 [56, 57] 。
为了研究P2Y2核苷酸受体对alpha分泌酶依赖性的淀粉样前体蛋白加工的影响, Cytoskeleton的羊抗人肌动蛋白抗体被用于蛋白质印迹 [58] 。
ICN Biomedicals的鼠源单克隆抗肌动蛋白抗体被用于LNCaP和PC-3细胞样品的蛋白质印迹 [59] 。
alpha和beta微管蛋白都一直被用作内参。
Santa Cruz Biotechnology的抗alpha微管蛋白抗体被用于表达有MT-I/II或MT-III的人类成纤维细胞GM00637 中 [60] ,以及用于研究Che-1在p21WAF1/Cip1激活中的作用 [61] 。 Santa Cruz Biotechnology的抗beta微管蛋白抗体被用于786-O细胞 [33] 和K562细胞 [62] 。为了研究RECQL4抗体的特异性,Santa Cruz的抗微管蛋白抗体(TU-02)被用于蛋白质印迹 [63] 。
在LNCaP、PC3M细胞中 [64] ,为了研究由泛素-蛋白酶体通路介导的结肠腺瘤样息肉病蛋白下调 [65] ,在表达有MuSK蛋白的COS细胞中 [66] ,在人类脐静脉血管内皮细胞中 [67] ,在T24和Hela细胞中 [68] ,Sigma的抗alpha微管蛋白抗体被用作内参。在人头颈部癌细胞株A253中 [69] ,在脊肌萎缩症(SMA)病人的成纤维细胞培养物中 [70] 以及MCF-7细胞、HCT116 p53+/+细胞、RKO细胞、p53野生型M7TS90细胞和p53缺失的M7TS90-E6细胞中 [71] ,使用了Sigma的单克隆抗beta微管蛋白抗体。Sigma的抗微管蛋白单克隆抗体(克隆DM1A)被用于胚胎和成人脑组织标本 [72] ,以及瞬时转染了载体、RACK1或突变型RACK1的NIH 3T3细胞中 [73] 。
BD PharMingen的抗beta微管蛋白抗体被用于来自于HeLa Tet-on细胞、HeLa Tet-on可诱导型FOXO4稳定细胞(15-14)以及稳定转染有野生型人源VHL表达质粒的BJAB细胞或RCC4细胞 [74] 的样品的蛋白质印迹。Oncogene Science的单克隆抗微管蛋白抗体也被用于Jurkat细胞 [75] 。
在文献 [76] 中,在小鼠的新皮层匀浆和N2a、ScN2a细胞中 [77] ,在人胎盘中 [78] ,在人胎儿肺实质中分离得到的上皮细胞中 [79] 以及球囊损伤后的雄性新西兰白兔中 [80] , EMD Millipore / Chemicon的抗GAPDH抗体均被用作对照。
为了确定在小鼠红系细胞中GAPDH的蛋白水平 [81] 以及在视网膜色素上皮细胞中 [29] , Abcam的单克隆抗GAPDH抗体被用于蛋白质印迹。
为了研究由心肌细胞PDGFR-beta信号通路介导的对压力超负荷小鼠心脏响应的调控 [82] , Santa Cruz Biotechnology的抗GAPDH抗体被用作内参。
AbDSerotec的鼠源抗GAPDH单克隆抗体在11种大鼠组织中被用于蛋白质印迹 [83] 。
在CD34+人类祖细胞、骨髓细胞和肿瘤细胞中,为了保证相同的上样量, HyTest的单克隆甘油醛-3-磷酸脱氢酶抗体(克隆6C5)被用于蛋白质印迹 [84] 。
同样也使用了来自于 Biodesign International [85] 、 Covance [86] 和Sigma Aldrich [87] 的抗体。
Santa Cruz Biotechnology HSC70抗体在分化中的造血干细胞中 [88] 和正常人成纤维细胞GM38中 [89] 被用作内参。 Stressgen Biotech抗HSP70抗体在P388D1细胞中被用于蛋白质印迹 [90] 。 Santa Cruz Biotechnology多克隆抗HSP90抗体在人类肥大细胞 [91] 和人源Hep3B细胞,以及大鼠FAO肝癌细胞中 [92] 被用作蛋白质印迹中的胞浆内参。
在人类HEK293细胞中为了保证相同的蛋白上样量, Santa Cruz Biotechnology特异性针对增殖细胞核抗原的抗体(PC-10)在蛋白质印迹中被用作内参 [93] 。在小鼠胚胎成纤维细胞中, DAKO的单克隆抗增殖细胞核抗原抗体(PC-10)在蛋白质印迹中被用作蛋白质上样量的参照 [94] 。
为了研究在乳腺癌细胞中雌激素受体alpha的调控 [95] ,为了研究MTA1和MAT1间的相互作用 [96] ,Sigma的抗黏着斑蛋白抗体在蛋白质印迹中被用作内参。
在很多情况下,有许多其它的蛋白靶点也一直被用作内参。这包括Arf1 [97] 、beta连环蛋白(beta-catenin) [98] 、钙联接蛋白(calnexin) [99] 、ERK-1 [19] 、ERK2 [100] 、HDAC1 [33] 、PDGFR alpha [91] 、转录因子IID [1] 、转铁蛋白受体 [2] 、核纤层蛋白A/C [45] 和肌球蛋白重链 [1] 、LHCP [3] ,以及Lhcb4 [4] 。
- Dittmer A, Dittmer J. Beta-actin is not a reliable loading control in Western blot analysis. Electrophoresis. 2006;27:2844-5 pubmed
- Olave I, Reck Peterson S, Crabtree G. Nuclear actin and actin-related proteins in chromatin remodeling. Annu Rev Biochem. 2002;71:755-81 pubmed
- Barber R, Harmer D, Coleman R, Clark B. GAPDH as a housekeeping gene: analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genomics. 2005;21:389-95 pubmed
- Yamaji R, Fujita K, Takahashi S, Yoneda H, Nagao K, Masuda W, et al. Hypoxia up-regulates glyceraldehyde-3-phosphate dehydrogenase in mouse brain capillary endothelial cells: involvement of Na+/Ca2+ exchanger. Biochim Biophys Acta. 2003;1593:269-76 pubmed
- Zhong H, Simons J. Direct comparison of GAPDH, beta-actin, cyclophilin, and 28S rRNA as internal standards for quantifying RNA levels under hypoxia. Biochem Biophys Res Commun. 1999;259:523-6 pubmed
- Schmitz H, Bereiter Hahn J. Glyceraldehyde-3-phosphate dehydrogenase associates with actin filaments in serum deprived NIH 3T3 cells only. Cell Biol Int. 2002;26:155-64 pubmed
- Yang Z, Barnes C, Kumar R. Human epidermal growth factor receptor 2 status modulates subcellular localization of and interaction with estrogen receptor alpha in breast cancer cells. Clin Cancer Res. 2004;10:3621-8 pubmed
- Talukder A, Mishra S, Mandal M, Balasenthil S, Mehta S, Sahin A, et al. MTA1 interacts with MAT1, a cyclin-dependent kinase-activating kinase complex ring finger factor, and regulates estrogen receptor transactivation functions. J Biol Chem. 2003;278:11676-85 pubmed
- Solan J, Fry M, TenBroek E, Lampe P. Connexin43 phosphorylation at S368 is acute during S and G2/M and in response to protein kinase C activation. J Cell Sci. 2003;116:2203-11 pubmed
- Ikari A, Nakano M, Kawano K, Suketa Y. Up-regulation of sodium-dependent glucose transporter by interaction with heat shock protein 70. J Biol Chem. 2002;277:33338-43 pubmed
- Anwar K, Fazal F, Malik A, Rahman A. RhoA/Rho-associated kinase pathway selectively regulates thrombin-induced intercellular adhesion molecule-1 expression in endothelial cells via activation of I kappa B kinase beta and phosphorylation of RelA/p65. J Immunol. 2004;173:6965-72 pubmed
- Chandrasekar B, Mummidi S, Claycomb W, Mestril R, Nemer M. Interleukin-18 is a pro-hypertrophic cytokine that acts through a phosphatidylinositol 3-kinase-phosphoinositide-dependent kinase-1-Akt-GATA4 signaling pathway in cardiomyocytes. J Biol Chem. 2005;280:4553-67 pubmed
- Yu Y, Davicioni E, Triche T, Merlino G. The homeoprotein six1 transcriptionally activates multiple protumorigenic genes but requires ezrin to promote metastasis. Cancer Res. 2006;66:1982-9 pubmed
- Granja A, Nogal M, Hurtado C, Vila V, Carrascosa A, Salas M, et al. The viral protein A238L inhibits cyclooxygenase-2 expression through a nuclear factor of activated T cell-dependent transactivation pathway. J Biol Chem. 2004;279:53736-46 pubmed
- Robinson White A, Hundley T, Shiferaw M, Bertherat J, Sandrini F, Stratakis C. Protein kinase-A activity in PRKAR1A-mutant cells, and regulation of mitogen-activated protein kinases ERK1/2. Hum Mol Genet. 2003;12:1475-84 pubmed
- Zhao J, Kong H, Li H, Huang B, Yang M, Zhu C, et al. IRF-8/interferon (IFN) consensus sequence-binding protein is involved in Toll-like receptor (TLR) signaling and contributes to the cross-talk between TLR and IFN-gamma signaling pathways. J Biol Chem. 2006;281:10073-80 pubmed
- Liu B, Faia L, Hu M, Nussenblatt R. Pro-angiogenic effect of IFNgamma is dependent on the PI3K/mTOR/translational pathway in human retinal pigmented epithelial cells. Mol Vis. 2010;16:184-93 pubmed
- Beaulieu J, Sotnikova T, Yao W, Kockeritz L, Woodgett J, Gainetdinov R, et al. Lithium antagonizes dopamine-dependent behaviors mediated by an AKT/glycogen synthase kinase 3 signaling cascade. Proc Natl Acad Sci U S A. 2004;101:5099-104 pubmed
- Ducker C, Upson J, French K, Smith C. Two N-myristoyltransferase isozymes play unique roles in protein myristoylation, proliferation, and apoptosis. Mol Cancer Res. 2005;3:463-76 pubmed
- Mauerer R, Ebert S, Langmann T. High glucose, unsaturated and saturated fatty acids differentially regulate expression of ATP-binding cassette transporters ABCA1 and ABCG1 in human macrophages. Exp Mol Med. 2009;41:126-32 pubmed
- Galban S, Martindale J, Mazan Mamczarz K, Lopez de Silanes I, Fan J, Wang W, et al. Influence of the RNA-binding protein HuR in pVHL-regulated p53 expression in renal carcinoma cells. Mol Cell Biol. 2003;23:7083-95 pubmed
- Nakamura S, Roth J, Mukhopadhyay T. Multiple lysine mutations in the C-terminal domain of p53 interfere with MDM2-dependent protein degradation and ubiquitination. Mol Cell Biol. 2000;20:9391-8 pubmed
- Ward I, Minn K, Chen J. UV-induced ataxia-telangiectasia-mutated and Rad3-related (ATR) activation requires replication stress. J Biol Chem. 2004;279:9677-80 pubmed
- Naderi S, Gutzkow K, Låhne H, Lefdal S, Ryves W, Harwood A, et al. cAMP-induced degradation of cyclin D3 through association with GSK-3beta. J Cell Sci. 2004;117:3769-83 pubmed
- Rosato R, Almenara J, Dai Y, Grant S. Simultaneous activation of the intrinsic and extrinsic pathways by histone deacetylase (HDAC) inhibitors and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) synergistically induces mitochondrial damage and apoptosis in human leukemia . Mol Cancer Ther. 2003;2:1273-84 pubmed
- Comi A, Hunt P, Vawter M, Pardo C, Becker K, Pevsner J. Increased fibronectin expression in sturge-weber syndrome fibroblasts and brain tissue. Pediatr Res. 2003;53:762-9 pubmed
- Kim K, Kim J, Lee J, Jin H, Lee S, Fisher D, et al. Nuclear factor of activated T cells c1 induces osteoclast-associated receptor gene expression during tumor necrosis factor-related activation-induced cytokine-mediated osteoclastogenesis. J Biol Chem. 2005;280:35209-16 pubmed
- McDowall A, Inwald D, Leitinger B, Jones A, Liesner R, Klein N, et al. A novel form of integrin dysfunction involving beta1, beta2, and beta3 integrins. J Clin Invest. 2003;111:51-60 pubmed
- Döppler H, Storz P, Li J, Comb M, Toker A. A phosphorylation state-specific antibody recognizes Hsp27, a novel substrate of protein kinase D. J Biol Chem. 2005;280:15013-9 pubmed
- Dentin R, Pégorier J, Benhamed F, Foufelle F, Ferre P, Fauveau V, et al. Hepatic glucokinase is required for the synergistic action of ChREBP and SREBP-1c on glycolytic and lipogenic gene expression. J Biol Chem. 2004;279:20314-26 pubmed
- Abid M, Schoots I, Spokes K, Wu S, Mawhinney C, Aird W. Vascular endothelial growth factor-mediated induction of manganese superoxide dismutase occurs through redox-dependent regulation of forkhead and IkappaB/NF-kappaB. J Biol Chem. 2004;279:44030-8 pubmed
- Aumais J, Williams S, Luo W, Nishino M, Caldwell K, Caldwell G, et al. Role for NudC, a dynein-associated nuclear movement protein, in mitosis and cytokinesis. J Cell Sci. 2003;116:1991-2003 pubmed
- Choi S, Kim M, Kang C, Bae S, Cho C, Soh J, et al. Activation of Bak and Bax through c-abl-protein kinase Cdelta-p38 MAPK signaling in response to ionizing radiation in human non-small cell lung cancer cells. J Biol Chem. 2006;281:7049-59 pubmed
- Griffin B, Moynagh P. Persistent interleukin-1beta signaling causes long term activation of NFkappaB in a promoter-specific manner in human glial cells. J Biol Chem. 2006;281:10316-26 pubmed
- Uemura N, Kajino T, Sanjo H, Sato S, Akira S, Matsumoto K, et al. TAK1 is a component of the Epstein-Barr virus LMP1 complex and is essential for activation of JNK but not of NF-kappaB. J Biol Chem. 2006;281:7863-72 pubmed
- Yin J, Kwon Y, Varshavsky A, Wang W. RECQL4, mutated in the Rothmund-Thomson and RAPADILINO syndromes, interacts with ubiquitin ligases UBR1 and UBR2 of the N-end rule pathway. Hum Mol Genet. 2004;13:2421-30 pubmed
- Ballestrero A, Nencioni A, Boy D, Rocco I, Garuti A, Mela G, et al. Tumor necrosis factor-related apoptosis-inducing ligand cooperates with anticancer drugs to overcome chemoresistance in antiapoptotic Bcl-2 family members expressing jurkat cells. Clin Cancer Res. 2004;10:1463-70 pubmed
- Piatelli M, Wardle C, Blois J, Doughty C, Schram B, Rothstein T, et al. Phosphatidylinositol 3-kinase-dependent mitogen-activated protein/extracellular signal-regulated kinase kinase 1/2 and NF-kappa B signaling pathways are required for B cell antigen receptor-mediated cyclin D2 induction in mature B cells. J Immunol. 2004;172:2753-62 pubmed
- Hocking D, Kowalski K. A cryptic fragment from fibronectin's III1 module localizes to lipid rafts and stimulates cell growth and contractility. J Cell Biol. 2002;158:175-84 pubmed
- Dai Y, Rahmani M, Corey S, Dent P, Grant S. A Bcr/Abl-independent, Lyn-dependent form of imatinib mesylate (STI-571) resistance is associated with altered expression of Bcl-2. J Biol Chem. 2004;279:34227-39 pubmed
- Chin L, Vavalle J, Li L. Staring, a novel E3 ubiquitin-protein ligase that targets syntaxin 1 for degradation. J Biol Chem. 2002;277:35071-9 pubmed
- Werner A, de Vries E, Tait S, Bontjer I, Borst J. Bcl-2 family member Bfl-1/A1 sequesters truncated bid to inhibit is collaboration with pro-apoptotic Bak or Bax. J Biol Chem. 2002;277:22781-8 pubmed
- Camden J, Schrader A, Camden R, Gonzalez F, Erb L, Seye C, et al. P2Y2 nucleotide receptors enhance alpha-secretase-dependent amyloid precursor protein processing. J Biol Chem. 2005;280:18696-702 pubmed
- Yang C, Lin H, Chen C, Yang Y, Tseng P, Rangnekar V, et al. Bcl-xL mediates a survival mechanism independent of the phosphoinositide 3-kinase/Akt pathway in prostate cancer cells. J Biol Chem. 2003;278:25872-8 pubmed
- Jeong H, Youn C, Cho H, Kim S, Kim M, Kim H, et al. Metallothionein-III prevents gamma-ray-induced 8-oxoguanine accumulation in normal and hOGG1-depleted cells. J Biol Chem. 2004;279:34138-49 pubmed
- Di Padova M, Bruno T, De Nicola F, Iezzi S, D Angelo C, Gallo R, et al. Che-1 arrests human colon carcinoma cell proliferation by displacing HDAC1 from the p21WAF1/CIP1 promoter. J Biol Chem. 2003;278:36496-504 pubmed
- Holstein S, Wohlford Lenane C, Hohl R. Consequences of mevalonate depletion. Differential transcriptional, translational, and post-translational up-regulation of Ras, Rap1a, RhoA, AND RhoB. J Biol Chem. 2002;277:10678-82 pubmed
- Petkovic M, Dietschy T, Freire R, Jiao R, Stagljar I. The human Rothmund-Thomson syndrome gene product, RECQL4, localizes to distinct nuclear foci that coincide with proteins involved in the maintenance of genome stability. J Cell Sci. 2005;118:4261-9 pubmed
- Rigas A, Ozanne D, Neal D, Robson C. The scaffolding protein RACK1 interacts with androgen receptor and promotes cross-talk through a protein kinase C signaling pathway. J Biol Chem. 2003;278:46087-93 pubmed
- Choi J, Park S, Costantini F, Jho E, Joo C. Adenomatous polyposis coli is down-regulated by the ubiquitin-proteasome pathway in a process facilitated by Axin. J Biol Chem. 2004;279:49188-98 pubmed
- Chevessier F, Faraut B, Ravel Chapuis A, Richard P, Gaudon K, Bauché S, et al. MUSK, a new target for mutations causing congenital myasthenic syndrome. Hum Mol Genet. 2004;13:3229-40 pubmed
- Hayward D, Clarke R, Faragher A, Pillai M, Hagan I, Fry A. The centrosomal kinase Nek2 displays elevated levels of protein expression in human breast cancer. Cancer Res. 2004;64:7370-6 pubmed
- Hendrickx N, Volanti C, Moens U, Seternes O, de Witte P, Vandenheede J, et al. Up-regulation of cyclooxygenase-2 and apoptosis resistance by p38 MAPK in hypericin-mediated photodynamic therapy of human cancer cells. J Biol Chem. 2003;278:52231-9 pubmed
- Brichta L, Hofmann Y, Hahnen E, Siebzehnrubl F, Raschke H, Blumcke I, et al. Valproic acid increases the SMN2 protein level: a well-known drug as a potential therapy for spinal muscular atrophy. Hum Mol Genet. 2003;12:2481-9 pubmed
- Longley D, Allen W, McDermott U, Wilson T, Latif T, Boyer J, et al. The roles of thymidylate synthase and p53 in regulating Fas-mediated apoptosis in response to antimetabolites. Clin Cancer Res. 2004;10:3562-71 pubmed
- Burgess H, Reiner O. Alternative splice variants of doublecortin-like kinase are differentially expressed and have different kinase activities. J Biol Chem. 2002;277:17696-705 pubmed
- Mamidipudi V, Zhang J, Lee K, Cartwright C. RACK1 regulates G1/S progression by suppressing Src kinase activity. Mol Cell Biol. 2004;24:6788-98 pubmed
- Tang T, Lasky L. The forkhead transcription factor FOXO4 induces the down-regulation of hypoxia-inducible factor 1 alpha by a von Hippel-Lindau protein-independent mechanism. J Biol Chem. 2003;278:30125-35 pubmed
- Harper N, Hughes M, MacFarlane M, Cohen G. Fas-associated death domain protein and caspase-8 are not recruited to the tumor necrosis factor receptor 1 signaling complex during tumor necrosis factor-induced apoptosis. J Biol Chem. 2003;278:25534-41 pubmed
- Zhang Z, Rehmann H, Price L, Riedl J, Bos J. AF6 negatively regulates Rap1-induced cell adhesion. J Biol Chem. 2005;280:33200-5 pubmed
- Hudak K, Parikh B, Di R, Baricevic M, Santana M, Seskar M, et al. Generation of pokeweed antiviral protein mutations in Saccharomyces cerevisiae: evidence that ribosome depurination is not sufficient for cytotoxicity. Nucleic Acids Res. 2004;32:4244-56 pubmed
- Ishikura N, Clever J, Bouzamondo Bernstein E, Samayoa E, Prusiner S, Huang E, et al. Notch-1 activation and dendritic atrophy in prion disease. Proc Natl Acad Sci U S A. 2005;102:886-91 pubmed
- Gellhaus A, Schmidt M, Dunk C, Lye S, Kimmig R, Winterhager E. Decreased expression of the angiogenic regulators CYR61 (CCN1) and NOV (CCN3) in human placenta is associated with pre-eclampsia. Mol Hum Reprod. 2006;12:389-99 pubmed
- Foster C, Zhang P, Gonzales L, Guttentag S. In vitro surfactant protein B deficiency inhibits lamellar body formation. Am J Respir Cell Mol Biol. 2003;29:259-66 pubmed
- Ju H, Nerurkar S, Sauermelch C, Olzinski A, Mirabile R, Zimmerman D, et al. Sustained activation of p38 mitogen-activated protein kinase contributes to the vascular response to injury. J Pharmacol Exp Ther. 2002;301:15-20 pubmed
- Zhou D, Pawlik K, Ren J, Sun C, Townes T. Differential binding of erythroid Krupple-like factor to embryonic/fetal globin gene promoters during development. J Biol Chem. 2006;281:16052-7 pubmed
- Guais A, Solhonne B, Melaine N, Guellaen G, Bulle F. Goliath, a ring-H2 mitochondrial protein, regulated by luteinizing hormone/human chorionic gonadotropin in rat leydig cells. Biol Reprod. 2004;70:204-13 pubmed
- Schag K, Schmidt S, Muller M, Weinschenk T, Appel S, Weck M, et al. Identification of C-met oncogene as a broadly expressed tumor-associated antigen recognized by cytotoxic T-lymphocytes. Clin Cancer Res. 2004;10:3658-66 pubmed
- Akagi S, Ichikawa H, Okada T, Sarai A, Sugimoto T, Morimoto H, et al. The critical role of SRC homology domain 2-containing tyrosine phosphatase-1 in recombinant human erythropoietin hyporesponsive anemia in chronic hemodialysis patients. J Am Soc Nephrol. 2004;15:3215-24 pubmed
- Ku M, Howard S, Ni W, Lagna G, Hata A. OAZ regulates bone morphogenetic protein signaling through Smad6 activation. J Biol Chem. 2006;281:5277-87 pubmed
- Plenchette S, Cathelin S, Rébé C, Launay S, Ladoire S, Sordet O, et al. Translocation of the inhibitor of apoptosis protein c-IAP1 from the nucleus to the Golgi in hematopoietic cells undergoing differentiation: a nuclear export signal-mediated event. Blood. 2004;104:2035-43 pubmed
- Zhu Z, Ramos J, Kampa K, Adimoolam S, Sirisawad M, Yu Z, et al. Control of ASPP2/(53BP2L) protein levels by proteasomal degradation modulates p53 apoptotic function. J Biol Chem. 2005;280:34473-80 pubmed
- Rusinol A, Thewke D, Liu J, Freeman N, Panini S, Sinensky M. AKT/protein kinase B regulation of BCL family members during oxysterol-induced apoptosis. J Biol Chem. 2004;279:1392-9 pubmed
- Qi X, Wildey G, Howe P. Evidence that Ser87 of BimEL is phosphorylated by Akt and regulates BimEL apoptotic function. J Biol Chem. 2006;281:813-23 pubmed
- Wang B, Liu K, Lin F, Lin W. A role for 14-3-3 tau in E2F1 stabilization and DNA damage-induced apoptosis. J Biol Chem. 2004;279:54140-52 pubmed
- Audebert M, Salles B, Calsou P. Involvement of poly(ADP-ribose) polymerase-1 and XRCC1/DNA ligase III in an alternative route for DNA double-strand breaks rejoining. J Biol Chem. 2004;279:55117-26 pubmed
- 来邦网
- 英文来邦
- 基因